Blog
Non-structural carbohydrates mediate seasonal water stress across Amazon forests
- Caroline Signori-Müller,
- Rafael S. Oliveira,
- David Galbraith
(Writers)
Plants rely on both newly assimilated carbon and stored reserves of non-structural carbohydrates (NSC) for growth and other physiological functions such as respiration, osmotic regulation and defence. As NSC stores reflect the balance of carbon supply via photosynthesis and its utilisation for plant metabolism, they are highly dynamic in time. NSC stored during periods when supply exceeds demand are thought to constitute an important buffer during periods of environmental stress when carbon demand outstrips supply. As a result of this, considerable attention has been paid to the potential role of stored NSC in mediating tree tolerance and survival under drought, during which stomatal conductance and assimilation rates are reduced to prevent water loss. Experimental studies on both temperate and tropical seedlings have indicated an important role of NSC in the physiological mechanism of mortality, as plants with higher NSC content had higher survivorship under drought. However, the extent to which NSC metabolism moderates tolerance to water deficit in adult trees over large geographical domains remains unclear.
An understanding of the functional role of NSC in response to water deficit is of particular importance for the Amazon rainforest, the Earth’s largest tropical forest region and a major terrestrial carbon sink, responsible for absorbing 5–10% of global anthropogenic CO2 emissions.
Over the last 15 years, the Amazon has been subject to three large-scale drought events and climate models project an intensification of drought risk over large parts of the Basin in the future. Recent modelling results suggest that NSC play an important role in regulating the impacts of drought on carbon fluxes in the Amazon by maintaining growth under water deficit. Moreover, observations of sustained stand-scale net primary productivity during the 2010 drought have also led to suggestions that Amazon trees deplete their NSC reserves during periods of water stress to prioritise growth. However, empirical studies of NSC dynamics in tropical forests are rare, being limited to a small number of sites and species. While a study in Panama found that NSC concentrations increased in the dry season, the only detailed community-level study in lowland Amazon forests, in a throughfall exclusion experiment in Eastern Amazonia, found that NSC reserves in trees subjected to long-term drought did not differ from those in unstressed trees. Yet Amazon forests vary greatly in climate, soils and plant life history strategies of which could potentially influence NSC dynamics and forest response to climate change. The scarcity of empirical data in Amazonia impedes understanding of the significance of NSC in modulating forest responses to water stress and thus limits current vegetation model development efforts to simulate drought impacts on tropical forests. To address this significant data gap, we conduct a large-scale sampling of NSC across Amazon forests, using fully standardised field and laboratory protocols, performing all NSC analyses in the same lab (see Methods). Such standardisation is critical as differences in sampling and laboratory extraction protocols can yield substantial variation in NSC estimates, obstructing meta-analysis and comparisons across studies.
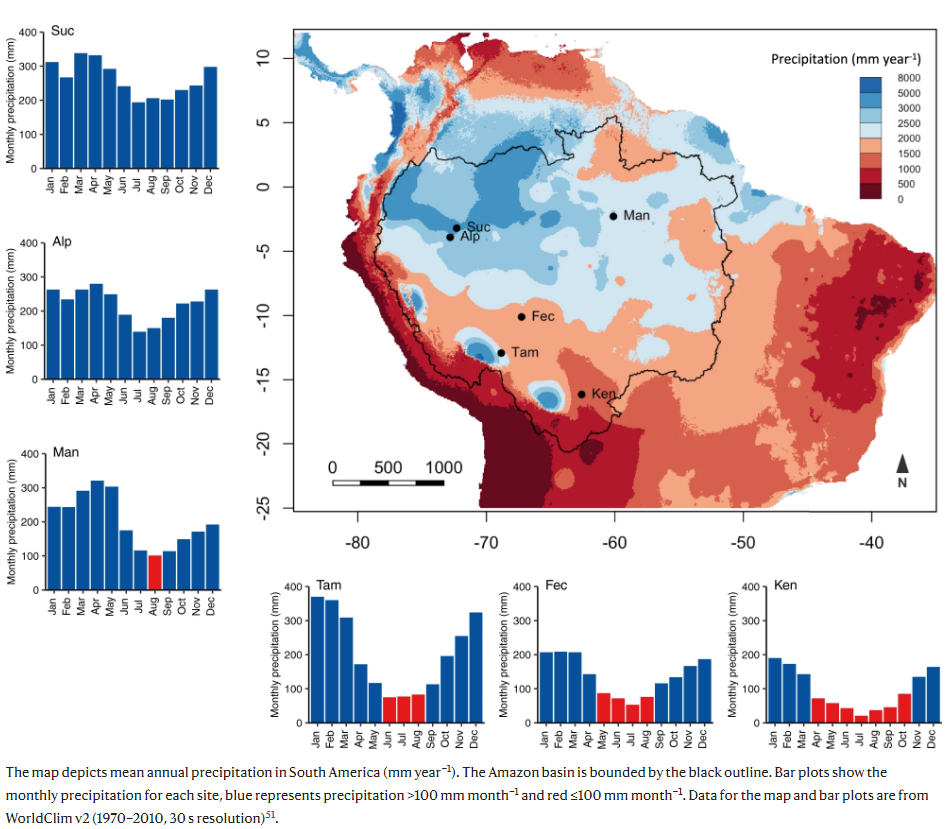
We analyse the concentration of NSC in leaves and branches of 82 canopy tree species in six sites across the Amazon Basin effectively spanning the entire Amazon gradient in mean annual precipitation (1167–3155 mm year−1; Fig. ) and seasonality (0–7 months with rainfall ≤100 mm month−1) and including one site (Man) that experienced an atypically strong drought event. We collect the plant material for NSC analyses in all sites during wet months (precipitation >100 mm month−1), hereafter referred to as the wet season. For the four sites with more marked seasonality (Ken, Fec, Man and Tam), we also collect plant material in the peak of the dry season, where monthly precipitation is ≤100 mm month−1 (Supplementary Fig. ). We focus not only on total NSC concentrations (NSCT) but also on the partitioning of NSC into its two major components: soluble sugars (i.e., oligosaccharides such as glucose, sucrose, fructose, etc.) and starch, as these fulfil distinct roles in plants. Soluble sugars (SS) provide an immediate energy substrate for respiration, defence, plant stress signalling, phloem transport and osmoregulation. Starch represents a transient or long-term energy store that plants can convert to SS for use when C demand exceeds supply. To characterise plant water status at the time of NSC sampling, we measure midday leaf water potential (ΨMD) in all sites during the dry season, and in the two sites without a climatological dry season (Alp and Suc). Community-level (mean value of all species in each site) ΨMD ranges from −0.62 ± 0.05 MPa (mean ± SE) in the ever-wet Suc and Alp sites to −2.18 ± 0.30 MPa in the ecotonal Ken site with the longest dry season.
